УДК 631.421.2:577.217
Применение нового молекулярного маркера и метода SDS-PAGE для установления взаимосвязи аллельного состава генов глютенинов с качественными показателями зерна тритикале
Пырсиков А. С.1, кандидат сельскохозяйственных наук
Сыксин С. В.1
Милюкова Н. А.1, кандидат биологических наук
Груздев И. В.1,2, кандидат биологических наук
Полховская Е. С.1
Гарибян Ц. С.1, кандидат технических наук
Соловьёв А. А.1,2, доктор биологических наук
1ФГБНУ «Всероссийский научно-исследовательский институт сельскохозяйственной биотехнологии»
127550, Россия, Москва, ул. Тимирязевская, д. 42
2ФГБУН «Главный ботанический сад им. Н. В. Цицина РАН»
127276, Россия, г. Москва, Ботаническая ул., д. 4
E-mail: andrey.pyrsikov@yandex.ru
Исследования выполнены в 2021–2022 годах в ФГБНУ «Всероссийский научно-исследовательский институт сельскохозяйственной биотехнологии» (г. Москва). В ходе работы был проведён скрининг коллекции из 26 образцов яровой гексаплоидной тритикале и яровой пшеницы с помощью классической ПЦР и SDS-PAGE с целью разработки новых эффективных молекулярно-генетических маркеров для идентификации аллелей генов высокомолекулярных глютенинов (локусы Glu-А1 и Glu-B1). Исследование амфидиплоидной культуры тритикале, объединяющей геномы пшеницы и ржи, биоинформатический поиск новых аллельных вариантов, кодирующих глютенины, открывает огромные возможности для работы с новыми маркерами этих генов и установления взаимосвязей между ними и показателями качества зерна. Содержание белка определяет питательную и кормовую ценность зерна и продуктов его переработки, а также технологические свойства зерновой массы. Белок тритикале характеризуется хорошо сбалансированным аминокислотным составом, занимая промежуточное положение между белками пшеницы и ржи по содержанию аспарагиновой кислоты, треонина, аланина, валина и тирозина. Идентификация субъединиц глютенинов и секалинов тритикале сопряжена с определёнными трудностями, так как они присутствуют в разных соотношениях и имеют сходную электрофоретическую подвижность. Молекулярный метод идентификации во многих случаях является более информативным, применяется в том числе и для характеристики генотипов тритикале по технологическим качествам зерна. Целью исследования была идентификация генов, кодирующих определённые субъединицы высокомолекулярных глютенинов (аллелей локусов Glu-A1 и Glu-B1) у сортов и сортообразцов тритикале, с помощью разработанных праймеров. В исследуемой коллекции сравнивали комбинации субъединиц высокомолекулярных глютенинов, определённые на основе ПЦР-маркирования и SDS-PAGE. В статье были идентифицированы наиболее перспективные с селекционной точки зрения образцы яровой тритикале. Применение методов SDS-PAGE, ПЦР-маркирование в сочетании с определением качественных показателей зерна тритикале позволило провести оценку генотипов по технологическим параметрам.
Ключевые слова: яровая тритикале; качество зерна; глютенины; молекулярные маркеры; ПЦР; SDS-PAGE.
Тритикале (× Triticosecale Wittmaсk.) — зерновая культура, представляющая собой амфидиплоид (аллополиплоид) пшеницы и ржи, в настоящее время она выделена в отдельный самостоятельный полиморфный ботанический род × Triticosecale.
Содержание белка в зерне тритикале варьируется от 12 до 25%, в среднем — около 15%, поскольку зависит от множества факторов: почвенно-климатических условий, генотипа, условий возделывания (агротехнических мероприятий), сроков посева и сбора урожая. В целом биохимический состав зерна тритикале является типичным для плода злаковых культур (Goutam et al., 2013; Обухова, Будашкина, 2014; Грабовец, Крохмаль, 2018; Абделькави и др., 2020).
Зерно тритикале более полноценно в пищевом и кормовом отношении в сравнении с зерном пшеницы вследствие более высокого содержания лизина (3,0 и 3,5% у пшеницы и тритикале соответственно). Содержание таких аминокислот, как гистидин, аргинин, изолейцин, лейцин, пролин, фенилаланин, также превышает родительские виды (Гольдварг и др., 2010; Goutam et al., 2013; Сокол, 2014; Энзекрей и др., 2020).
В зерне тритикале, как и большинства зерновых культур, выделяют легкорастворимые белки (альбумины и глобулины), которые составляют 15–20% от общего содержания белков в зерне, и белки клейковины — 80–85% (Dubois et al., 2016; Kilic et al., 2017; Dubois et al., 2018). При этом эндосперм тритикале содержит около 26–28% альбуминов, 7–8% глобулинов, 25–26% проламинов и 18–20% глютелинов.
Традиционно именно субъединичный состав высокомолекулярных глютенинов становится предметом масштабных исследований с помощью электрофореза в полиакриламидном геле (SDS-PAGE и/или A-PAGE) для их идентификации и установления влияния на хлебопекарные и кормовые качества как пшеницы, так и тритикале, которая имеет всего четыре гена высокомолекулярных субъединиц глютенина, как правило, одна-три субъединицы синтезируются в конкретном сорте (Дивашук и др., 2010). Гены глютенина (HMW) кодируются в локусах Glu-A1 и Glu-B1 в длинных плечах хромосом 1A и 1B. Каждый локус включает два тесно сцепленных гена, кодирующих два типа субъединиц глютенина, обозначенные как x-тип и y-тип. Соответствующие гены HMW секалина ржи кодируются в локусе Glu-R1, также обозначенном как Sec-3, который расположен в длинном плече хромосомы 1R (Дивашук и др., 2007). Локус Glu-D1 пшеницы положительно влияет на качество хлеба, соответственно, при его отсутствии у тритикале снижается общее количество глютена в зерне и, соответственно, качество выпечки. Хотя существует генетическая изменчивость содержания глютена, даже самое высокое содержание белка клейковины в тритикале по-прежнему на 10–15% ниже, чем у пшеницы (Payne, 1984; Payne et al., 1987; Amiour et al., 2002; Geng et al., 2014; Kilic et al., 2017).
Запасные белки тритикале (HMW и LMW глютенин и субъединицы секалина), идентифицированные с помощью SDS-PAGE, показывают значительно более низкий диапазон, чем у пшеницы. Наличие у тритикале запасных белков как пшеницы, так и ржи, которые присутствуют в разных количествах и, кроме того, часто характеризуются сходной электрофоретической подвижностью, в значительной степени мешает идентификации индивидуальных субъединиц высокомолекулярных глютенинов с помощью SDS-PAGE.
Поэтому более полную информацию о хлебопекарных качествах образца можно получить при совместном использовании метода электрофореза в полиакриламидном геле и молекулярного метода идентификации генотипов тритикале, несущих разные комбинации аллелей глютенинов локусов Glu-A1 и Glu-B1 (Amiour et al., 2002; Salmanowicz, Dylewicz, 2007; Kilic et al., 2017; Liu et al., 2017; Шумный, Обухова, 2018).
В последнее время молекулярный метод идентификации генотипов тритикале в связи с её хлебопекарными качествами получает всё более широкое применение. Это касается как уже упомянутых локусов Glu-A1 и Glu-B1, так и других генов, так или иначе связанных с хорошими хлебопекарными характеристиками, таких как ген wbm (wheat bread making), локализованный на хромосоме 7А (Furtado et al., 2015; Liu et al., 2017; Kirov et al., 2019).
Целью исследования являлась идентификация генов, кодирующих определённые субъединицы высокомолекулярных глютенинов (аллелей локусов Glu-A1 и Glu-B1) у сортов и сортообразцов тритикале, с помощью разработанных праймеров. Сравнивали комбинации субъединиц высокомолекулярных глютенинов в исследуемой коллекции тритикале, определённые на основе ПЦР-маркирования и SDS-PAGE.
Методика исследований. Исследования выполнены в 2021–2022 годах в ФГБНУ «Всероссийский научно-исследовательский институт сельскохозяйственной биотехнологии» (г. Москва). В исследовании использовали 24 образца яровой тритикале (Гребешок, Ярило, С95, С238, С245, С259, Л8665, Л8666, 08844, V17-150, П2-16-20, П1-13-5-2, П1-13-5-13, ПРАГ 551, 131/7, 131/714 (Россия), Памяти Мережко, Ульяна, Лана (Беларусь), Укро, Хлебодар украинский (Украина), Sandro (Швейцария), Dublet, Legalo (Польша) и два образца мягкой яровой пшеницы Chinese Spring (Китай), Лада (Россия)) различного географического происхождения из коллекции лаборатории маркерной и геномной селекции растений ФГБНУ ВНИИСБ.
Выделение ДНК осуществляли из свежих проростков растений с помощью набора «ДНК-Экстран 3» (ОАО «Синтол», Россия). Измерение концентрации ДНК осуществляли с использованием NanoPhotometr (Implen, Германия). Дизайн праймеров осуществляли на основании биоинформатических данных (BLAST, UGENE).
Для проведения ПЦР использовали следующую смесь: ПЦР-буфер для Taq polymerase pH=8,4, dNTP 250 µM, MgCl2 3 mM, Taq polymerase 3-6 U на реакцию, 60 нг ДНК. Условия проведения ПЦР модифицировали для разработанной пары праймеров Glu 396/352: F: CAACCTCTCCACAGCAGC, R: CTTGTCCTGACTGTTGCG, температура отжига — 65oC, ожидаемый размер ампликонов — 244 пары нуклеотидов (п.н.) (Glu Bx-17), 335 п.н. (Glu Bx-14), 352 п.н. (Glu Bx-7 и Glu Bx-13) и 396 п.н. (Glu Bx-6).
Электрофорез ДНК проводили в 1х ТАЕ буфере с использованием 2%-го агарозного геля при режиме: 3 В/см — 10 минут; 3,5 В/см — 110 минут. Результаты детектировали с помощью УФ-камеры Gel Doc XR+ (Bio-Rad, США). Электрофорез белков проводили в пластинках полиакриламидного геля (концентрирующий: T=5%; C=2,67%; разделяющий: T=12,8%; C=0,99%) размером 20×20×0,1 см, в течение 19,5 ч, при силе тока 16 мА, в камере для вертикального электрофореза PROTEAN® II xi производства компании BioRad. Электродный буфер трис-глициновый. После электрофореза гели фиксировали в уксусном алкоголе и окрашивали Coomassie Brilliant Blue R-250.
Образцы фрагментов, которые визуально отличались на электрофореграмме по размеру от ожидаемого размера продукта амплификации, были выделены из геля для дальнейшего лигирования в плазмиду и трансформации E. coli, с последующей постановкой ПЦР колоний и секвенированием.
Секвенирование ПЦР-продуктов выполнено компанией «Синтол» (ОАО «Синтол», Москва). По результатам секвенирования были получены последовательности, которые были проанализированы с помощью BLAST-анализа на соответствие с последовательностями аллелям генов глютенинов. Определение качественных показателей зерна тритикале проводили с помощью прибора «Инфра ЛЮМ ФТ-12» (Люмекс, Россия). Масса одной пробы составляла не менее 100 г. Результаты измерений анализировали с помощью программы «СпектраЛЮМ/Про®» (Люмекс, Россия).
Результаты исследований. SDS-PAGE анализ. Образцы белка 26 образцов изучаемой коллекции были проанализированы по составу субъединиц высокомолекулярных глютенинов с помощью одномерного электрофореза (SDS-PAGE). Два образца мягкой яровой пшеницы — Chinese Spring и Лада — использовали как стандарт. На рис. 1 представлена электрофореграмма сорта Гребешок, различающаяся по аллелям в двух локусах (Glu-A1 и Glu-B1) и локусе ржи (Glu-R1).
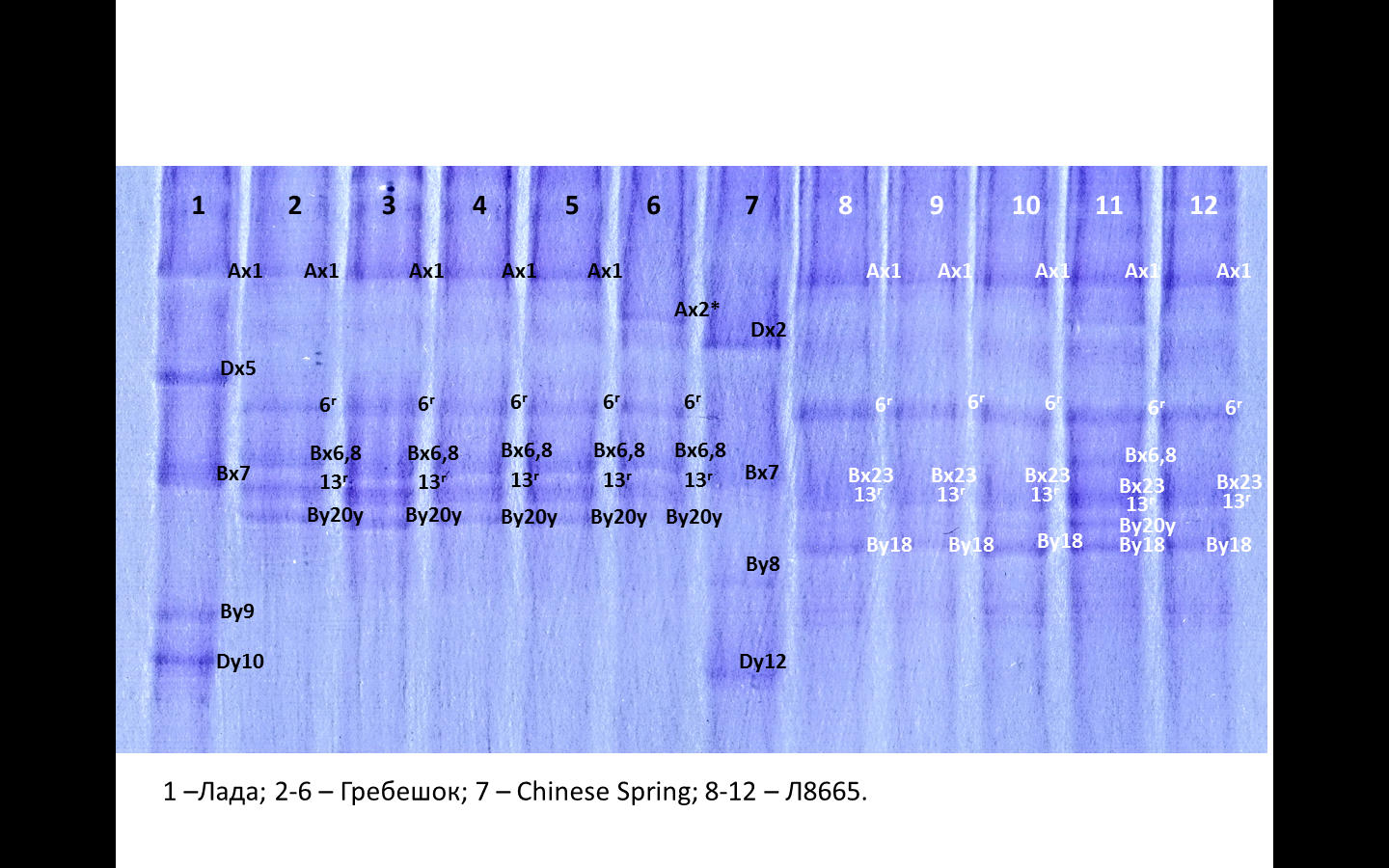
Рис. 1. Идентификация высокомолекулярных субъединиц глютенинов (локусы Glu-A1 и Glu-B1) у образца Гребешок из изучаемой коллекции:
1 — Лада (Ax1/Bx7+By9/Dx5+Dy10); 2 — Гребешок (Ax1/Bx6,8+By20y/6r+13r); 3 — Гребешок (Ax1/Bx6,8+By20y/6r+13r); 3 — Гребешок (Ax1/Bx6,8+By20y/6r+13r); 4 — Гребешок (Ax1/Bx6,8+By20y/6r+13r); 5 — Гребешок (Ax2*/Bx6,8+By20y/6r+13r); 6 — Chinese Spring (AxN/Bx7+By8/Dx2+Dy12)
Большинство сортообразцов, по данным анализа, были гетерогенными. Комбинации аллелей (субъединиц) различны у образцов коллекции и имеют разную частоту встречаемости (табл. 1).
ПЦР-анализ генов глютенинов. Данные, полученные в результате ПЦР-анализа с разработанными ген- или аллель-специфичными ПЦР маркерами, которые использовались в этом исследовании для идентификации субъединиц высокомолекулярных глютенинов, в дальнейшем были сопоставлены с данными SDS-PAGE анализа.
На рис. 2 представлены результаты электрофореза при использовании пары праймеров Glu 396/352. По наличию у образцов яровой тритикале фрагментов ожидаемого размера сделан вывод о присутствии соответствующих аллелей локусов Glu-A1 и Glu-B1. Итоговые результаты представлены в табл. 1.
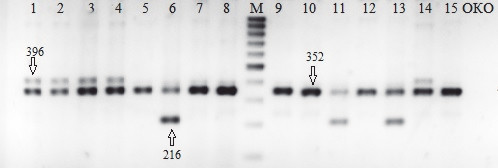
Рис. 2. Продукты амплификации с использованием праймера Glu 396/352.
Образцы: 1 — П1-13-5-13 (а); 2 — П1-13-5-13 (б); 3 — П1-13-5-2 (а); 4 — П1-13-5-2 (б); 5 — Гребешок (а); 6 — Гребешок (б); 7 — Ульяна (а); 8 — Ульяна (б); М — маркер молекулярного веса 100bp+; 9 — С259 (а); 10 — С259 (б); 11 — Л8665 (а); 12 — Л8665 (б); 13 — Хлебодар (а); 14 — Хлебодар (б); 15 — Chines Spring; ОКО — отрицательный контрольный образец
В результате проведённой работы у некоторых проанализированных образцов с праймером Glu 396/352 были обнаружены фрагменты, длина которых отличалась от ожидаемой и составила 216 и 420 п.н.
Образцы фрагментов, которые визуально отличались на электрофореграмме по размеру от ожидаемого размера продукта амплификации, были выделены из геля для дальнейшего лигирования в плазмиду и трансформации E. coli, с последующей постановкой ПЦР колоний и секвенированием. По результатам секвенирования было установлено, что фрагмент 216 п.н. соответствует варианту аллелей Glu Bx-7 и Glu-Ax, а фрагмент 420 п.н. — варианту аллеля Glu Ax.
1. Комбинации высокомолекулярных субъединиц глютенинов по результатам SDS PAGE и ПЦР-анализа
Образец | Комбинация высокомолекулярных субъединиц глютенинов и секалинов (по результатам SDS PAGE) | Комбинация высокомолекулярных субъединиц глютенинов (по итогам ПЦР-анализа) | |||
геном А (Glu-A1) | геном B (Glu-B1) | геном R (Glu-R1) | геном А (Glu-A1) | геном B (Glu-B1) | |
Chinese Spring | N | 7+8 | D/2+12 | – | 7, 8 |
Лада | 1 | 7+9 | Dx5+Dy10 | – | – |
Укро | 40% — 1; 60% — N | 80% — 7+8; 20% — 6,8+20y | 80% — 1r+4r; 20% — 6r+13r | – | 6, 7 |
Гребешок | 80% — 1; 20% — 2* | 6,8+20y | 6r+13r | – | 6, 7 |
Ульяна | 25% — 1; 75% — N | 7+8 | 75% — 1r+4r; 25% — 6r+13r | – | 6, 7 |
Ярило | 2* | 23+18 | 6r+13r | Ax | 7 |
Памяти Мережко | 2* | 6,8+20y | 6r+13r | Ax | 7 |
Sandro | 1 | 6,8+20y | 6r+13r | Ax | 7 |
Лана | N | 7+18 | 1r+4r | Ax | 7 |
Dublet | 2* | 33% — 6,8+20y; 67% — 7+18 | 6r+13r | Ax | 7, 17 7 |
Legalo | 2* | 7+8 | 2r+6,5r | Ax | 7 |
131/7 | N | 7+9 | 60% — 2r+6,5r; 40% — 1r+4r | Ax | 14 |
131/714 | N | 7+9 | 1r+4r | – | 6, 7 |
C 259 | 80% — 1; 20% — N | 80% — 6,8+20y; 20% — 6,8+20y/7+8 | 80% — 6r+13r; 20% — 1r+4r | – | 7 |
C 238 | 80% — 2*; 20% — N | 6,8+20y | 6r+13r | Ax | 7 |
Л 8665 | 1 | 80% — 23+18; 20% — 23+18/6,8+20y | 6r+13r | Ax | 7 |
C 95 | 7,7% — N; 7,7% — 2*; 84,6% — 1 | 69,2% — 7+18; 7,7% — 7+8; 7,7% — 23+18; 15,4% — 6,8+20y. | 7,7% — 6r+13r; 92,3% — 1r+4r | Ax | 7 |
08844 | 36,4% — N; 54,5% — 2*; 9,1% — 1 | 81,8% — 6,8+20y; 9,1% — 23+18/6,8+20y; 9,1% — 7+9/6,8+20y. | 18,2% — 1r+4r; 18,2% — 6r+13r; 9,1% — 1r+4r/6r+13r; 54,5% — 5,8r/6r+13r | Ax | 7 |
V17-150 | 66,7% — 2*; 33,3% — N | 7+9 | 6r+13r | Ax | 7 |
П2-16-20 | 90% — 2*; 10% — N | 90% — 6,8+20y; 10% — 7+9 | 90% — 6r+13r; 10% — 2r+6,5r. | – | 6, 7 |
ПРАГ 551 | 1 | 6,8+20y | 6r+13r | – | 6, 7 |
Л 8666 | 50% — 2*; 50% — N | 50% — 6,8+20y; 50% — 7+9 | 6r+13r | — | 7 |
С245 | 2* | 7+18 | 2r+6,5r | — | 7 |
П1-13-5-13 | 66,6% — N; 16,7% — 2*; 16,7% — 1 | 50% — 6,8+20y; 16,7% — 13+16; 16,7% — 7+18; 16,7% — 7+8 | 50% — 6r+13r; 50% — 1r+4r. | – | 6, 7 |
П1-13-5-2 | 57,1% — N; 28,6% — 2*; 14,3% — 1 | 57,1% — 7+18; 28,6% — 7+8; 14,3% — 6,8+20y | 6r+13r | Ax | 7 |
Хлебодар | N | 50% — 7+8; 50% — 7+9 | 1r+4r | Ax | 7 |
Разнообразие аллельных вариантов локусов Glu-A1 и Glu-B1 у сортов тритикале очевидно. Локус Glu-B1 более полиморфен, чем локус Glu-A1. Наиболее распространёнными типами являются субъединицы 6, 7, 13 и 17 х-типа. Кроме того, субъединица 7 имеет, по крайней мере, одну вариацию с несколько более высокой молекулярной массой и более выраженной полосой в SDS-PAGE. Разнообразие локуса Glu-B1 по субъединицам у-типа ещё больше: 8, 9, 14, 16, 18, 20, 21 и 25. В нашем исследовании в исследуемых образцах ярового тритикале мы выделили пять аллелей локуса Glu-B1: Glu-B1b, ответственный за синтез комбинации 7+8 субъединиц; Glu-B1c, отвечающий за синтез субъединиц 7+9; Glu-B1IV — 23+18; Glu-B1r — 7+18; Glu-B1s — 6,8+20y. Субъединица Bx7, кодируемая Glu-B1, широко распространена среди сортов пшеницы и тритикале, оказывает положительное влияние на технологические свойства, но обычно связана с аллельными субъединицами By8 или By9, что затрудняет оценку влияния одной Bx7 как компонента качества зерна. В результате идентификации высокомолекулярных субъединиц глютена методом SDS-PAGE у сортов Ульяна и Легало обнаружен аллель Glu-B1b (7+8). Кроме того, этот аллель обнаружен у 80% исследованных растений сорта Укро и у 50% растений сорта Хлебодар. Аллель Glu-B1b также был идентифицирован в 20% образцов С259; однако эти растения были гетерозиготными. Кроме того, линии Р13-5-2 и Р13-5-13 характеризовались наличием этого аллеля у 28,6 и 16,7% растений соответственно. Наличие аллеля Glu-B1b у данных образцов может косвенно указывать на высокий хлебопекарный потенциал указанных выше образцов. Аллель Glu-B1c (7+9) выявлен у линий 131/7, В17-150 и 131/714, у 50% образцов сорта Хлебодар, у 50% образцов Л8666, у 10% растений в линии Р2-16-20 и у 9,1% растений линии 08844, которые являются гетерозиготными. Комбинация субъединиц Bx7 и By9 обычно связана с высокой прочностью теста. Аллель Glu-B1r, определяющая синтез высокомолекулярных субъединиц 7+18, репрезентативен только у тритикале, выявлен у сорта Лана, а также у сорта Дублет. Наличие этого аллеля характеризует линию С245 и 57,1% растений линии Р13-5-2. Аллель s локуса Glu-B1 наиболее часто встречается в генотипах изученных образцов ярового гексаплоидного тритикале. Он характеризуется субъединицами 6,8 + 20у и идентифицирован только у тритикале. Этот аллель был идентифицирован у сортов Гребешок, Памяти Мережко и Сандро, а также в образцах С238, L8665 и PRAG 551. В образце 08844 он обнаружен у всех изученных растений; однако 18,2% растений были гетерозиготами. Аналогичная картина наблюдалась и у образца С259. Гетерогенными образцами с аллелем Glu-B1s были Укро (20%), Дублет (33%), Л8666 (50%), Р13-5-13 (50%) и Р13-5-2 (14,3%).
Для того чтобы подтвердить взаимосвязь обнаруженных фрагментов, отвечающих за наличие определённых субъединиц высокомолекулярных глютенинов, с технологическими качествами зерна тритикале, было проведено определение качественных показателей зерна имеющихся образцов с помощью прибора «Инфра ЛЮМ ФТ-12» (Люмекс, Россия), результаты которого представлены в табл. 2.
2. Качественные показатели зерна пшеницы и тритикале изучаемой коллекции
Образец | Влажность, % | Белок, % | Клейковина, % | ИДК, ед. | Крахмал, % | Условная крахмалистость, % | Растяжимость, см |
Chinese Spring | 10,96 | 15,40 | 27,44 | – | 54,23 | 53,27 | – |
Лада | 9,82 | 14,85 | 24,74 | – | 57,17 | 60,33 | – |
Укро | 6,74 | 13,26 | 15,19 | 85,1 | 67,04 | 56,40 | 13,6 |
Гребешок | 6,88 | 13,49 | 15,81 | 87,5 | 67,35 | 55,34 | 14,7 |
Ульяна | 6,72 | 13,19 | 14,74 | 82,6 | 68,32 | 56,84 | 14,7 |
Ярило | 6,69 | 15,54 | 18,16 | 88,9 | 65,73 | 54,39 | 16,3 |
Памяти Мережко | 6,61 | 15,72 | 19,59 | 92,8 | 64,68 | 53,60 | 18,7 |
Sandro | 6,53 | 15,26 | 20,25 | 85,0 | 65,77 | 53,96 | 20,8 |
Лана | 6,69 | 14,49 | 15,84 | 88,6 | 67,37 | 55,65 | 16,9 |
Dublet | 6,43 | 14,38 | 15,69 | 85,7 | 67,36 | 56,47 | 17,7 |
Legalo | 6,88 | 13,43 | 17,61 | 80,9 | 67,95 | 56,44 | 10,7 |
131/7 | 7,59 | 12,70 | 20,09 | 51,7 | 65,06 | 53,87 | 19,7 |
131/714 | 6,52 | 14,85 | 17,35 | 91,9 | 65,21 | 54,64 | 13,0 |
С259 | 6,74 | 16,01 | 22,39 | 96,4 | 64,31 | 52,18 | 19,7 |
С238 | 6,67 | 14,27 | 16,44 | 89,3 | 66,55 | 55,59 | 13,4 |
Л8665 | 6,33 | 14,61 | 18,79 | 82,9 | 67,03 | 54,88 | 21,7 |
С95 | 7,46 | 13,48 | 14,36 | 67,5 | 66,49 | 57,58 | 9,6 |
О8844 | 7,80 | 14,49 | 30,41 | 57,5 | 63,45 | 51,89 | 26,1 |
V17-150 | 7,82 | 12,60 | 18,00 | 55,8 ед | 65,02 | 54,67 | 17,9 |
П2-16-20 | 6,54 | 15,35 | 21,80 | 88,3 | 65,97 | 54,30 | 23,2 |
ПРАГ 551 | 7,21 | 10,90 | 15,18 | 51,1 | 68,78 | 56,68 | 18,9 |
Л8666 | 8,65 | 13,76 | 22,88 | 55,0 | 63,28 | 51,74 | 23,1 |
С245 | 7,16 | 12,50 | 16,16 | 57,8 | 67,07 | 56,98 | 19,2 |
П13-5-13 | 8,25 | 13,11 | 17,93 | 63,4 | 64,63 | 53,19 | 20,5 |
П2-13-5-2 | 6,60 | 14,95 | 19,60 | 88,2 | 65,88 | 53,82 | 22,3 |
Хлебодар | 6,33 | 13,92 | 18,03 | 81,5 | 67,27 | 55,98 | 21,9 |
Как известно, технологические свойства зерна определяются высоким содержанием белка, клейковины и хорошей растяжимостью. Глютенины как неотъемлемый компонент клейковины во многом определяют кормовые и хлебопекарные качества зерна. По данным ранее проведённых исследований (Salmanowicz, 2007; Kilic, 2017), селекционный отбор в ранних поколениях на субъединицы Ax1, Ax2 и Bx7 и идентификация соответствующих аллелей локусов Glu-A1 и Glu-B1 на основе молекулярно-генетического анализа могут оказать положительное влияние на повышение технологических показателей качества зерна яровой тритикале.
На основании этого среди изученных образцов яровой тритикале следует выделить пять образцов как наиболее перспективных, а именно Гребешок, С95, 08844, П1-13-5-13 и П1-13-5-2 (табл. 1). По данным табл. 2, клейковина образцов Гребешок, С95, П1-13-5-13 является хорошей по показателю ИДК (ГОСТ Р 54478-2011).
Заключение. Таким образом, в настоящем исследовании коллекция образцов и сортообразцов тритикале проанализирована по технологическим параметрам. Разработаны и протестированы аллель-специфические маркеры для идентификации субъединиц глютенина сортов и сортообразцов тритикале. Сравнительные данные SDS-PAGE, ПЦР-анализа и инструментального метода определения показателей качества зерна подтвердили высокую эффективность исследуемых маркеров. Таким образом, предложенные в данном исследовании методы при совместном применении позволили четко идентифицировать субъединицы глютенинов и выделить наиболее ценные генотипы.
Исследование выполнено при частичной поддержке гранта Президента Российской Федерации № МК-3235.2021.5 и государственного задания № FGUM-2022-0005.
Литература
- Стабильность и пластичность генотипов яровой тритикале по урожайности и качеству зерна / Р. Н. Абделькави, О. А. Щуклина, О. И. Ермоленко, А. А. Соловьёв // Аграрный научный журнал. — 2020. — № 4. — С.4–9.
- Озимое тритикале — ценная зерновая культура / Б. А. Гольдварг, В. Г Гриценко, Л. И. Бораева, В. Я. Ковтуненко // Тритикале: материалы международной практической конференции «Роль тритикале в стабилизации и увеличении производства зерна и кормов» и секции тритикале отделения растениеводства РАСХН. — Ростов-на-Дону: ДЗНИИСХ, 2010. — С.284–287.
- Грабовец А. И. Тритикале: монография / А. И. Грабовец, А. В. Крохмаль. — Ростов-на Дону: ООО «Идательство «Юг», 2018. — 240 с.
- Дивашук М. Г. Влияние отбора по фенотипическим признакам на хромосомную конституцию яровой тритикале / М. Г. Дивашук, А. А. Соловьёв, Г. И. Карлов // Генетика. — 2010. — Т. 46. — № 3. — С.383–388.
- Дивашук М. Г. Использование микросателлитных маркеров для идентификации пшенично-ржаной транслокации у гексаплоидной тритикале / М. Г. Дивашук, Г. И. Карлов, А. А. Соловьёв // Известия Тимирязевской сельскохозяйственной академии. — 2007. — № 1. — С.61–65.
- Обухова Л. В. Корреляционный анализ зависимости силы муки от запасных белков пшеницы / Л. В. Обухова, Е. Б. Будашкина // Вавиловский журнал генетики и селекции. — 2014. — Т. 18. — № 4/1. — С.807–811.
- Сокол Н. В. Тритикале — культура хлебная / Н. В. Сокол. — Palmarium Academic Publishing, 2014. — 145 с.
- Шумный В. К. Состав высокомолекулярных субъединиц глютенина у сортов и перспективных линий мягкой пшеницы / В. К. Шумный, Л. В. Обухова // Генетика. — 2018. — С.316–325.
- Связь генов проламинов с хлебопекарными качествами зерна тритикале / Е. С. Энзекрей, Н. А. Милюкова, А. С. Пырсиков, С. В. Сыксин // Сборник материалов Международной научно-практической конференции (27 ноября 2020 г., г. Краснодар). Кубанский филиал ФГБНУ «ФНЦ пищевых систем им. В. М. Горбатова» РАН. — Краснодар, 2020. — С.165–167.
- Энзекрей Е. С. Влияние метеорологических условий и азотных удобрений на биологическую урожайность яровой тритикале сорта Тимирязевская 42 / Е. С. Энзекрей, О. А. Щуклина, С. В. Завгородний // Зерновое хозяйство России. — 2021. — № 2 (74). — С.88–93.
- Diversity of seven glutenin and secalin loci within triticale cultivars grownin Europe / N. Amiour, A. Bouguennec, C. Marcoz, P. Sourdille, M. Bourgoin, D. Khelifi, G. Branlard // Euphytica. — 2002. — No. 123. — P.295–305.
- Genetic and environmental factors affecting the expression of α-gliadin canonical epitopes involved in celiac disease in a wide collection of spelt (Triticum aestivum ssp. spelta) cultivars and landraces / B. Dubois, P. Bertin, L. Hautier, Y. Muhovski, E. Escarnot, D. Mingeot // Plant Biology. — 2018. — Vol. 18 — P.262.
- Dubois B. Molecular diversity of α-gliadin expressed genes in genetically contrasted spelt (Triticum aestivum ssp. spelta) accessions and comparison with bread wheat (T. aestivum ssp. aestivum) and related diploid Triticum and Aegilops species / B. Dubois, P. Bertin, D. Mingeot // Mol. Breeding. — 2016. — Vol. 36. — P.152–155. Doi: 10.1007/s11032-016-0569.
- A novel highly differentially expressed gene in wheat endosperm associated with bread quality / A. Furtado, P. C. Bundock, P. Banks, G. Fox, X. Yin, R. Henry // Sci. Rep. — 2015. — P.10–46.
- Geng Y. Expression of wheat high molecular weight glutenin subunit 1Bx is affected by large insertions and deletions located in the upstream flanking sequences / Y. Geng, B. Pang, C. Hao // PloS One. — 2014. — Vol. 9. — No 8. — P.25–31.
- Biotechnological approaches for grain quality improvement in wheat: present status and future possibilities / U. Goutam, S. Kukreja, R. Tiwari, R. K. Gupta, B. B. Dholakia, R. Yadav // Australian Journal of Crop Science. — 2013. — Vol. 7. — No. 4. — P.469.
- Sources of the highly expressed wheat bread making (wbm) gene in CIMMYT spring wheat germplasm and its effect on processing and bread-making quality/ C. Guzmán, Y. Xiao, J. Crossa, H. González-Santoyo, J. Huerta, R. Singh, S. Dreisigacker // Euphytica. — 2016. — Vol. 209. — No. 3. — P.689–692.
- Screening bread wheat genotypes for high molecular weight glutenin subunits and some quality parameters / H. Kilic, T. Sanal, I. Erdemci, K. Karaca // J. Agric. Sci. Technol. — 2017. — Vol. 19. — P.1393–1404.
- Analysis of Wheat Bread-Making Gene (wbm) Evolution and Occurrence in Triticale Collection Reveal Origin via Interspecific Introgression into Chromosome 7AL / I. Kirov, A. Pirsikov, N. Milyukova, M. Dudnikov, M. Kolenkov, I. Gruzdev, S. Siksin, L. Khrustaleva, G. Karlov, A. Soloviev // Agronomy. — Basel, Switzerland, 2019. — P.1–9.
- Potential for marker-assisted simultaneous improvement of grain and biomass yield in triticale / W. Liu, H. P. Maurer, W. L. Leiser, M. R. Tucker, S. Weissmann, V. Hahn, T. Würschum // Bioenergy Research. — 2017. — Vol. 10 (2). — P.449–455.
- Payne P. I. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality / P. I. Payne // Annual Review of Plant Physiology. — 1987. — Vol. 38. — No. 1. — P.141–153.
- Wheat storage proteins: their genetics and their potential for manipulation by plant breeding / P. I. Payne, L. M. Holt, E. A. Jackson, C. N. Law // Philosophical Transactions of the Royal Society of London. Biological Sciences. — 1984. — Vol. 304. — No. 1120. — P.359–371.
- Salmanowicz B. P. Identification and characterization of high molecular weight glutenin genes in Polish triticale cultivars by PCR-generated DNA markers / B. P. Salmanowicz, M. Dylewicz // J. Appl. Genet. — 2007. — Vol. 48. — Р.347–357.
Correlation analyses between the allelic composition of glutenin genes and the quality of triticale grain via new molecular marker and SDS-PAGE
Pyrsikov A. S.1, PhD Agr. Sc.
Syksin S. V.1
Milyukova N. A.1, PhD Biol. Sc.
Gruzdev I. V.1,2, PhD Biol. Sc.
Polkhovskaya E. S.1
Garibyan Ts. S.1, PhD Techn. Sc.
Soloviev A. A.1,2, Dr. Biol. Sc.
1The All-Russia Research Institute of Agricultural Biotechnology
127550, Russia, Moscow, Timiryazevskaya str., 42
2Leading Botanical Garden n. a. N. V. Tsitsin of the Russian Academy of Sciences
127276, Russia, Moscow, Botanicheskaya str., 4
E-mail: andrey.pyrsikov@yandex.ru
This investigation took place at the All-Russia Research Institute of Agricultural Biotechnology (Moscow) in 2021–2022. Twenty-six genotypes of spring hexaploid triticale and wheat were analyzed via PCR and SDS-PAGE to design new effective genetic markers identifying the alleles of high molecular weight (HMW) glutenin genes (Glu-А1 and Glu-B1 loci). Triticale as an amphidiploid crop comprising both wheat and rye genomes reveals new bioinformatic insights into finding new glutenin-coding alleles and correlations between genetic markers and grain quality. Protein content defines the nutritional and forage values of grain and grain products as well as the technological properties of grain mass. Triticale protein has balanced amino acid composition. In aspartic acid, threonine, alanine, valine and tyrosine it stands between the proteins of wheat and rye. The identification of triticale glutenin and secalin subunits has its pitfalls since these proteins are presented in various ratios and have similar electrophoretic mobility. Molecular identification is more efficient method and can be used to distinguish triticale genotypes according to the technological properties of their grain. The aim of this research was to identify genes coding HMW glutenin subunits (alleles of the Glu-A1 and Glu-B1 loci) of triticale genotypes using the primers designed. The combinations of HMW glutenin subunits were compared that were identified via PCR and SDS-PAGE. This article reports on the most promising genotypes of spring triticale to be used in breeding projects. SDS-PAGE and PCR together with the traits of grain quality enabled the genotype testing by technological properties.
Keywords: spring triticale; grain quality; glutenin; molecular marker; PCR; SDS-PAGE.
References
- Stabilnost i plastichnost genotipov yarovoy tritikale po urozhaynosti i kachestvu zerna / R. N. Abdelkavi, O. A. Shchuklina, O. I. Ermolenko, A. A. Solovev // Agrarnyy nauchnyy zhurnal. — 2020. — No. 4. — P.4–9.
- Ozimoe tritikale — tsennaya zernovaya kultura / B. A. Goldvarg, V. G Gritsenko, L. I. Boraeva, V. Ya. Kovtunenko // Tritikale: materialy mezhdunarodnoy prakticheskoy konferentsii “Rol tritikale v stabilizatsii i uvelichenii proizvodstva zerna i kormov” i sektsii tritikale otdeleniya rastenievodstva RASKhN. — Rostov-na-Donu: DZNIISKh, 2010. — P.284–287.
- Grabovets A. I. Tritikale: monografiya / A. I. Grabovets, A. V. Krokhmal. — Rostov-na Donu: OOO “Idatelstvo “Yug”, 2018. — 240 p.
- Divashuk M. G. Vliyanie otbora po fenotipicheskim priznakam na khromosomnuyu konstitutsiyu yarovoy tritikale / M. G. Divashuk, A. A. Solovev, G. I. Karlov // Genetika. — 2010. — Vol. 46. — No. 3. — P.383–388.
- Divashuk M. G. Ispolzovanie mikrosatellitnykh markerov dlya identifikatsii pshenichno-rzhanoy translokatsii u geksaploidnoy tritikale / M. G. Divashuk, G. I. Karlov, A. A. Solovev // Izvestiya Timiryazevskoy selskokhozyaystvennoy akademii. — 2007. — No. 1. — P.61–65.
- Obukhova L. V. Korrelyatsionnyy analiz zavisimosti sily muki ot zapasnykh belkov pshenitsy / L. V. Obukhova, E. B. Budashkina // Vavilovskiy zhurnal genetiki i selektsii. — 2014. — Vol. 18. — No. 4/1. — P.807–811.
- Sokol N. V. Tritikale — kultura khlebnaya / N. V. Sokol. — Palmarium Academic Publishing, 2014. — 145 p.
- Shumnyy V. K. Sostav vysokomolekulyarnykh subedinits glyutenina u sortov i perspektivnykh liniy myagkoy pshenitsy / V. K. Shumnyy, L. V. Obukhova // Genetika. — 2018. — P.316–325.
- Svyaz genov prolaminov s khlebopekarnymi kachestvami zerna tritikale / E. S. Enzekrey, N. A. Milyukova, A. S. Pyrsikov, S. V. Syksin // Sbornik materialov Mezhdunarodnoy nauchno-prakticheskoy konferentsii (27 noyabrya 2020 g., g. Krasnodar). Kubanskiy filial FGBNU “FNTs pishchevykh sistem im. V. M. Gorbatova” RAN. — Krasnodar, 2020. — P.165–167.
- Enzekrey E. S. Vliyanie meteorologicheskikh usloviy i azotnykh udobreniy na biologicheskuyu urozhaynost yarovoy tritikale sorta Timiryazevskaya 42 / E. S. Enzekrey, O. A. Shchuklina, S. V. Zavgorodniy // Zernovoe khozyaystvo Rossii. — 2021. — No. 2 (74). — P.88–93.
- Diversity of seven glutenin and secalin loci within triticale cultivars grownin Europe / N. Amiour, A. Bouguennec, C. Marcoz, P. Sourdille, M. Bourgoin, D. Khelifi, G. Branlard // Euphytica. — 2002. — No. 123. — P.295–305.
- Genetic and environmental factors affecting the expression of α-gliadin canonical epitopes involved in celiac disease in a wide collection of spelt (Triticum aestivum ssp. spelta) cultivars and landraces / B. Dubois, P. Bertin, L. Hautier, Y. Muhovski, E. Escarnot, D. Mingeot // Plant Biology. — 2018. — Vol. 18 — P.262.
- Dubois B. Molecular diversity of α-gliadin expressed genes in genetically contrasted spelt (Triticum aestivum ssp. spelta) accessions and comparison with bread wheat (T. aestivum ssp. aestivum) and related diploid Triticum and Aegilops species / B. Dubois, P. Bertin, D. Mingeot // Mol. Breeding. — 2016. — Vol. 36. — P.152–155. Doi: 10.1007/s11032-016-0569.
- A novel highly differentially expressed gene in wheat endosperm associated with bread quality / A. Furtado, P. C. Bundock, P. Banks, G. Fox, X. Yin, R. Henry // Sci. Rep. — 2015. — P.10–46.
- Geng Y. Expression of wheat high molecular weight glutenin subunit 1Bx is affected by large insertions and deletions located in the upstream flanking sequences / Y. Geng, B. Pang, C. Hao // PloS One. — 2014. — Vol. 9. — No 8. — P.25–31.
- Biotechnological approaches for grain quality improvement in wheat: present status and future possibilities / U. Goutam, S. Kukreja, R. Tiwari, R. K. Gupta, B. B. Dholakia, R. Yadav // Australian Journal of Crop Science. — 2013. — Vol. 7. — No. 4. — P.469.
- Sources of the highly expressed wheat bread making (wbm) gene in CIMMYT spring wheat germplasm and its effect on processing and bread-making quality/ C. Guzmán, Y. Xiao, J. Crossa, H. González-Santoyo, J. Huerta, R. Singh, S. Dreisigacker // Euphytica. — 2016. — Vol. 209. — No. 3. — P.689–692.
- Screening bread wheat genotypes for high molecular weight glutenin subunits and some quality parameters / H. Kilic, T. Sanal, I. Erdemci, K. Karaca // J. Agric. Sci. Technol. — 2017. — Vol. 19. — P.1393–1404.
- Analysis of Wheat Bread-Making Gene (wbm) Evolution and Occurrence in Triticale Collection Reveal Origin via Interspecific Introgression into Chromosome 7AL / I. Kirov, A. Pirsikov, N. Milyukova, M. Dudnikov, M. Kolenkov, I. Gruzdev, S. Siksin, L. Khrustaleva, G. Karlov, A. Soloviev // Agronomy. — Basel, Switzerland, 2019. — P.1–9.
- Potential for marker-assisted simultaneous improvement of grain and biomass yield in triticale / W. Liu, H. P. Maurer, W. L. Leiser, M. R. Tucker, S. Weissmann, V. Hahn, T. Würschum // Bioenergy Research. — 2017. — Vol. 10 (2). — P.449–455.
- Payne P. I. Genetics of wheat storage proteins and the effect of allelic variation on bread-making quality / P. I. Payne // Annual Review of Plant Physiology. — 1987. — Vol. 38. — No. 1. — P.141–153.
- Wheat storage proteins: their genetics and their potential for manipulation by plant breeding / P. I. Payne, L. M. Holt, E. A. Jackson, C. N. Law // Philosophical Transactions of the Royal Society of London. Biological Sciences. — 1984. — Vol. 304. — No. 1120. — P.359–371.
- Salmanowicz B. P. Identification and characterization of high molecular weight glutenin genes in Polish triticale cultivars by PCR-generated DNA markers / B. P. Salmanowicz, M. Dylewicz // J. Appl. Genet. — 2007. — Vol. 48. — Р.347–357.