УДК 638.19:581.162.32:577.175.24
Привлекательность левзеи для медоносной пчелы
Авдеев Н. В., кандидат биологических наук
ФГБНУ «Пермский НИИСХ» – филиал ПФИЦ УрО РАН
614532, Россия, Пермский край, Пермский р-н, с. Лобаново, ул. Культуры, д. 12
E-mail: permbee@yandex.ru
Исследование проводилось в ФГБНУ «Пермский НИИСХ» на популяциях левзеи сафлоровидной и медоносных пчёл Apis mellifera mellifera L. в 2019 году. Целью исследования являлась оценка возможности опыления левзеи сафлоровидной медоносными пчёлами в условиях Пермского края. Была проанализирована доля пчёл, опыляющих исследуемый вид, при использовании нектара и цветочной пыльцы. При выборе нектара важным фактором является содержание и соотношение в нём сахаров. Левзея сафлоровидная — перспективное лекарственное и кормовое растение, возделывание которого главным образом ограничено отсутствием качественных семян. В период максимального цветения левзеи проводили отлавливание пчёл, возвращающихся в улей с ношами собранной цветочной пыльцы (обножками). При идентификации пыльцевых зёрен с помощью специальных справочников и микропрепаратов определяли вид, род, семейство или тип растения. По результатам пыльцевого анализа обнаружено, что летом 2019 года в Пермском крае левзея не выделяла нектар в количестве, доступном для сбора пчёлами (частота встречаемости пыльцевых зёрен левзеи в мёде составила менее 1%). Тем не менее пчёлы активно собирали пыльцу левзеи, доля пчёл с обножками левзеи составляла 22,5%, что является высоким показателем, учитывая площадь посадки левзеи (0,6 га) и наличие других энтомофильных растений в пределах продуктивного радиуса пчёл (3 км). Скорее всего, в пыльце левзеи присутствуют экдистероиды, обуславливающие её высокую привлекательность для медоносной пчелы. Использование медоносной пчелы может стать важным элементом технологии получения качественных семян левзеи сафлоровидной.
Ключевые слова: пчела медоносная, левзея сафлоровидная, пыльцевой анализ, опыление, экдистероиды.
Левзея сафлоровидная Rhaponticum carthamoides (Willd.) Iljin. — эндемик субальпийских лугов Алтай-Саянского региона, перспективное лекарственное и кормовое растение. Главной причиной, ограничивающей её широкое возделывание, является отсутствие качественных семян (Алябьева, 2005). В то же время семенная продуктивность представляет собой один из важнейших показателей адаптации вида к конкретным условиям обитания.
Литературные данные по семенной продуктивности левзеи часто основаны на разных методах её оценки и показывают значительные различия, обусловленные рядом факторов, в том числе её зависимостью от опыления (Головко, 1996; Карусевич, 2016; Рубан, 2013; Тимофеев, 2008). Зачастую приходят к заключению о необходимости искусственного опыления культуры, предотвращающего потери урожая семян в результате их недоразвития (Карусевич, 2016). Антэкология левзеи и возможность использования медоносной пчелы для её опыления практически не изучены.
Левзея по комплексу морфолого-физиологических характеристик является типичным представителем энтомофильных растений. Венчик фиолетово-лилового цвета, цветки собраны в крупные соцветия в виде плотной, почти округлой верхушечной корзинки диаметром 4–8 см. Цветки обоеполые, пятичленные, трубчатые, нектароносная ткань находится в основании столбика. Отмечена высокая мёдопродуктивность сплошных массивов левзеи, достигающая 100 кг/га (Бурмистров, 1992).
Особенностью левзеи является высокое содержание экдистероидов (главным образом 20-гидроксиэкдизона) как в вегетативных, так и в генеративных органах растения. Концентрация таких соединений может достигать 5,7% (Тимофеев, 1998). 20-гидроксиэкдизон является гормоном линьки насекомых, инициирующим метаморфозы в концентрации 10–8–10–9 М (Ахрем, 1989). Считается, что накопление экдистероидов в концентрации 10–5 М является результатом адаптации растений к насекомым-фитофагам. Такие соединения обладают репеллентным действием, способны нарушать развитие и вызывать гибель фитофагов (Дайнен, 1998; Уфимцев, 2009а; 2009б). Однако не для всех насекомых экдистероиды обладают инсектицидным действием. Более того, предполагается, что для насекомых-фитофагов (Oxythyrea funesta, Potosia cuprea metallica, Trichius fasciatus), поражающих левзею, привлекательность богатых экдистероидами репродуктивных органов обусловлена необходимостью получения фитоэкдистероидов в качестве источника стероидов для пластического обмена (Тимофеев, 2009).
Стерины — незаменимые для пчёл вещества, получаемые с пыльцой. Пыльца содержит различные фитостерины (С28, С29) и незначительные количества холестерина (С27) (Rasmont, 2005; Svoboda, 1983; Vanderplanck, 2014; Vanderplanck, 2018). В то же время медоносная пчела не может деалкилировать фитостерины в положении С-24 в 27-углеводные соединения холестеринового ряда (Herbert, 1980; Svoboda, 1983). Наряду с 20-гидроксиэкдизоном (С27) в качестве гормона линьки пчёлы используют макистерон А (С28) или макистерон С (С29) (Feldlaufer, 1986; Svoboda, 1982).
Возможности метаболизма стеринов определяют развитие потомства и пищевое поведение пчелиных (Rasmont, 2005; Vanderplanck, 2014; Vanderplanck, 2018). Например, земляничное дерево (Arbutus unedo) является основным кормовым видом для шмеля земляного в южной Франции в осенний период. Но пыльца этого вида богата β-ситостеролом, и медоносная пчела не собирает его пыльцу, хотя посещает цветы для сбора нектара (Rasmont, 2005). В опытах по искусственному кормлению микросемьи шмеля земляного пыльцой разных видов обнаружено, что пыльца бодяка (Cirsium) оказывает угнетающее действие на насекомых, особенно на количество и массу выращиваемых личинок. Эта пыльца характеризовалась наличием довольно редких δ7-стеринов. Интересно, что одним из побочных эффектов стеринов пыльцы бодяка было увеличение запасания углеводного корма (Vanderplanck, 2018).
Имеются сообщения (Пестов, 2017) о наличии экдистеронов (количественных данных нет) в нектаре трёх видов рода серпуха (Serratula), также обладающих высоким содержанием экдистероидов. При этом серпуха венценосная (S. coronata), содержащая помимо 20-гидроксиэкдизона экдизон и инокостерон, имеет консортивные связи с большим количеством видов пчелиных. В опытах по скармливанию семьям медоносной пчелы экдистеронового экстракта в концентрации 2×10–11 М показано его стимулирующее влияние на сбор мёда (Тимофеев, 1996).
Пчёлы могут собирать пыльцу, нектар или и то, и другое. Соотношение пчёл, работающих на сборе пыльцы и нектара, определяется генетически (Hunt, 1995; Page, 1995), но может существенно варьироваться в зависимости от количества открытого расплода (Keller et al., 2005) и интенсивности взятка (Полищук, 1984). При выборе нектара существенным фактором является содержание и соотношение в нём сахаров как для медоносной пчелы (Зауралов, 1980; 1981), и для других насекомых-опылителей (Abrol, 1991; Bogdanov, 1996). Привлекательность пыльцы обусловлена липидами трифины (оболочки пыльцевого зерна): стирольным соединением, ненасыщенной жирной кислотой, ненасыщенным С18-алканом или каким-то другим нейтральным липидом (Dabson, 1988; Johanson, 1977). При этом перечисленные липидные соединения должны обладать определённой летучестью, поскольку при мобилизации пчёл-новичков на сбор пыльцы они оценивают качество обножки дистанционно, без трофического контакта, необходимого при вербовке на сбор нектара (Фриш, 1980).
Целью исследования являлось изучение возможности использования медоносной пчелы для опыления левзеи сафлоровидной в условиях Пермского края. Была поставлена задача оценить долю пчёл, опыляющих исследуемый вид, при использовании нектара в качестве пищевого стимула и цветочной пыльцы.
Методика исследований. Исследование проводилось в ФГБНУ «Пермский НИИСХ» в 2019 году на семенном травостое левзеи сафлоровидной 10-го года жизни и медоносных пчёлах аборигенной среднерусской породы (расы) Apis mellifera mellifera L. Учётная площадь — 0,6 га. Перед цветением левзеи из семьи пчёл были максимально полно изъяты рамки с мёдом и поставлена надставка с пустыми соторамками. После окончания цветения собранный в надставке мёд был использован для пыльцевого анализа с целью установления ботанического происхождения. Определение частоты встречаемости пыльцевых зёрен в мёде проводили в соответствии с ГОСТ 31769-2012. В период максимального цветения левзеи отлавливали пчёл, возвращающихся в улей с ношами собранной цветочной пыльцы (обножками). Проведён анализ 250 пчёл с обножками. При идентификации пыльцевых зёрен с помощью специальных справочников определяли вид, род, семейство или тип растения (Бурмистров, Никитина, 1992; Карпович, 2015). Для сравнения также использовались собственные препараты собранной вручную пыльцы левзеи (рис. 1).
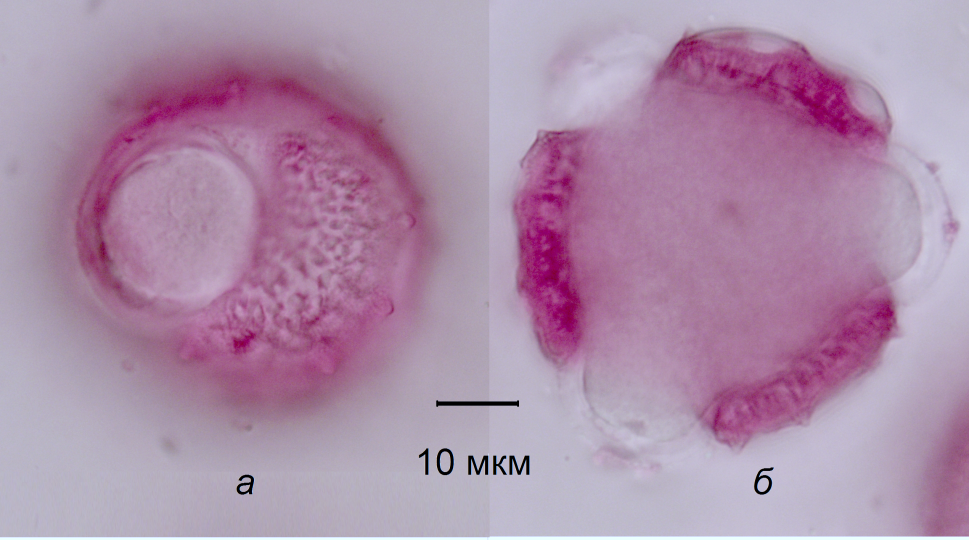
Рис. 1. Микрофотография пыльцевого зерна левзеи сафлоровидной (окраска фуксином):
а — экваториальная проекция; б — полярная проекция
Результаты исследований. Медоносная пчела интенсивно посещала левзею, многократно перелетая с соцветия на соцветие. Доля пыльцевых обножек, собранных пчёлами с левзеи, составила 22,5% (табл. 1). Это очень хороший результат, учитывая соотношение площади посадки левзеи и площади кормового участка пчёл в радиусе продуктивного лёта пчёл (3 км, 0,6 га/2828 га). При этом пчёлы игнорировали богатый пыльцой и нектаром одуванчик лекарственный, массово произрастающий в непосредственной близости от левзеи.
1. Ботаническое происхождение пыльцы
Ботаническое происхождение пыльцевых зёрен обножек | Доля пчёл с обножкой указанного ботанического происхождения, % |
Galega orientalis Lam. | 67,2 |
Rhaponticum carthamoides (Willd.) Iljin | 22,5 |
Salex sps. | 9,1 |
Несколько видов растений без доминирования одного из них | 1,2 |
При сборе пыльцы пчёлы проявляют «цветочное постоянство», посещая только один вид растения за один вылет (Гайдак, 1969). Исключение составляют пчёлы-разведчицы, собирающие пыльцу одновременно с нескольких видов растений без явного предпочтения. При сборе нектара «цветочное постоянство» менее выражено, поскольку в медовом зобике возвращающейся в улей пчелы могут содержаться пыльцевые зёрна нескольких видов растений, при этом пыльца одного из видов, как правило, доминирует (Авдеев, 2006).
При сборе пыльцы пчёлы эффективно посещали три вида растений: козлятник восточный, левзею сафлоровидную и один из видов ивы (табл. 1). По результатам пыльцевого анализа собранного мёда (табл. 2) с уверенностью можно сказать, что при сборе нектара пчёлы посещали козлятник восточный, один из видов ивы и малину обыкновенную. Наличие в мёде других пыльцевых зёрен (бодяка полевого) можно объяснить перемещением запасов прошлогоднего мёда с гнездовых рамок, случайным попаданием (пыльца ветроопыляемых злаков) и разведывательной активностью пчёл.
2. Результаты пыльцевого анализа мёда
Ботаническое происхождение пыльцевых зёрен, встречающихся в мёде | Частота встречаемости пыльцевых зёрен, (х ± ∆) % |
Galega orientalis Lam. | 37,9 ± 8,0 |
Salix sps. | 33,2 ± 7,0 |
Rubus idaeus L. | 14,7 ± 3,1 |
Myosotis sps. | 4,1 ± 0,9 |
Lupinus sps. | 2,7 ± 0,6 |
Papaveraceae | 1,5 ± 0,3 |
Trifolium repens L. | 2,0 ± 0,4 |
Filipendula sps. | 1,0 ± 0,2 |
Crucifereae | 1,0 ± 0,2 |
Taraxacum L. тип. | Менее 1 |
Rhaponticum carthamoides (Willd.) Iljin | Менее 1 |
Vicia L. тип | Менее 1 |
Prunus L. тип | Менее 1 |
Acer sps. | Менее 1 |
Cirsium arvense (L.) Scop. | Менее 1 |
Gramineae | Менее 1 |
Количество смешанных пыльцевых обножек, собираемых пчёлами-разведчицами, составило 1,2%, что соответствует литературным данным (Гайдак, 1969). Небольшое количество видов растений (по три вида при сборе как нектара, так и пыльцы), одновременно посещаемых для продуктивного сбора нектара и пыльцы, характерно для расы Apis mellifera mellifera L. (Авдеев, 2006; Жеребкин, Миронова, 1976).
Пыльцевые зёрна левзеи встречались в мёде, но их доля составляла менее 1%, что можно объяснить попаданием их в мёд с тел пчёл в улье или разведывательной активностью пчёл. Вероятно, в условиях 2019 года выделение нектара было недостаточным для наполнения трубки венчика до уровня, доступного для медоносной пчелы.
Заключение. Таким образом, даже при отсутствии пищевого стимула в виде нектара пчёлы активно посещали соцветия левзеи только для сбора пыльцы. Следовательно, использование медоносной пчелы может стать важным элементом агротехнологии получения качественных семян левзеи сафлоровидной.
Обнаруженная высокая привлекательность пыльцы левзеи для медоносной пчелы, возможно, обусловлена содержанием в ней экдистероидов.
Литература
- Авдеев Н. В. Особенности трофической специализации разных экологических групп евро-сибирской пчелы Apis mellifera mellifera L.: автореф. дис. … канд. биол. наук: 03.00.16, 06.02.04. — М., 2006. — 108 с.
- Алябьева Г. Н. Охрана и рациональное использование эндемичных растений Кузнецко-Салаирского нагорья / Г. Н. Алябьева, С. В. Овсянникова // Современные наукоёмкие технологии. — 2005. — № 5. — С.51–53.
- Ахрем А. А. Экдистероиды: химия и биологическая активность / А. А. Ахрем, Н. В. Ковганко. — Минск: Наука и техника, 1989. — 327 с.
- Бурмистров А. Н. Медоносные растения и их пыльца / А. Н. Бурмистров, В. А. Никитина. — М.: Росагропромиздат, 1990. — 192 с.
- Гайдак М. Г. Жизнедеятельность медоносных пчёл / М. Г. Гайдак // Пчела и улей. — М.: Колос, 1969. — С.80–148.
- Рапонтик сафлоровидный в культуре на Европейском Северо-Востоке (эколого-физиологическое исследование) / Т. К. Головко, Е. В. Гармаш, С. В. Куренкова, Г. Н. Табаленкова, Ю. М. Фролов. — Сыктывкар: Коми научный центр УрО РАН, 1996. — 140 с.
- Дайнен Л. Стратегия оценки роли фитоэкдистероидов как детеррентов по отношению к беспозвоночным фитофагам / Л. Дайнен // Физиология растений. — 1998. — Т. 45. — № 3. — С.347–359.
- Жеребкин М. В. О некоторых особенностях пыльцесобирательной деятельности пчёл разных пород / М. В. Жеребкин, Р. К. Миронова // Вестник. — 1976. — № 25.— С.3–61.
- Зауралов О. А. Растение и нектар. Образование и выделение нектара / О. А. Зауралов. — Саратов: Изд-во Саратовского университета, 1985. — 180 с.
- Зауралов О. А. Флороспециализация у медоносных пчёл / О. А. Зауралов // Журнал общей биологии. — 1981. — Т. 42. — № 6. — С.844–856.
- Атлас пыльцевых зёрен (Pollen atlas) / И. В. Карпович, Е. С. Дребезгина, Е. А. Еловикова, Г. И. Леготкина, Е. Н. Зубова, Р. З. Кузяев, Р. Х. Хисматуллин. — Екатеринбург: Уральский рабочий, 2015. — 318 с.
- Карусевич А. А. Разработка методики культивирования и изучения особенностей развития левзеи сафлоровидной в Витебской области / А. А. Карусевич, Г. Н. Осочук // Вестник фармации. — 2016. — № 4 (74). — С.38–44.
- Разнообразие антофильных насекомых и биохимические особенности нектара интродуцентов рода Serratula / С. В. Пестов, Н. И. Филиппов, К. Г. Уфимцев, В. В. Володин, С. О. Володина, А. Г. Донцов, И. В. Бешлей // Теоретическая и прикладная экология. — 2017. — № 3. — С.91–96.
- Полищук В. П. Сбор пыльцы в период медосбора / В. П. Полищук // Пчеловодство. — 1984. — № 11. — С.12–13.
- Рубан Г. А. Особенности семенной репродукции левзеи сафлоровидной / Г. А. Рубан, К. С. Зайнуллина // Аграрная наука Евро-Северо-Востока. — 2013. — № 4 (35). — С.22–25.
- Тимофеев Н. П. Распределение 20-гидроксиэкдизона в структуре биомассы надземной части Rhaponticum carthamoides (Willd.) Iljin в условиях агроценоза (Республика Коми) / Н. П. Тимофеев, В. В. Володин, Ю. М. Фролов // Растительные ресурсы. — 1998. — Т. 34. — № 3. — С.63–69.
- Тимофеев Н. П. Семенная репродукция и повреждаемость вредителями растений с гормональной активностью насекомых / Н. П. Тимофеев // Эколого-популяционный анализ полезных растений: интродукция, воспроизводство, использование. — Сыктывкар: Изд-во Коми НЦ УрО РАН, 2008. — С.192–194.
- Тимофеев Н. П. Экологическое взаимодействие агропопуляции экдистероидсодержащих растений с насекомыми-фитофагами. Сообщение 2. Изменчивость состава фитоэкдистероидов в аргоценозе и их роль в поражаемости растений вредителями / Н. П. Тимофеев // Сибирский экологический журнал. — 2009. — № 6. — С.829–842.
- Тимофеев Н. П. Эффект малых доз экдистероидов в пчеловодстве / Н. П. Тимофеев // Материалы IV международной научно-производственной конференции «Селекция, экология, технология возделывания и переработки нетрадиционных растений». — Симферополь, 1996. — С.231–232.
- Уфимцев К. Г. Фитоэкдистероиды — детерренты насекомых-фитофагов / К. Г. Уфимцев, Т. И. Ширшова, В. В. Володин. — Екатеринбург: Изд-во Коми НЦ УрО РАН, 2009. — 89 с.
- Уфимцев К. Г. Фитоэкдистероиды как детерренты насекомых-фитофагов: действие растения серпухи венценосной Serratula coronata L., продуцента экдистероидов, на египетскую хлопковую совку Spodoptera littorali (Lepidoptera: Noctuida) / К. Г. Уфимцев, Т. И. Ширшова, В. В. Володин // Успехи современной биологии. — 2009. — Т. 129. — № 3. — С.1–15.
- Фриш К. Из жизни пчёл / К. Фриш. — М.: Мир, 1980. — 216 с.
- Abrol D. P. Foraging strategies of honeybees and solitary bees as determined by nectar sugar components / D. P. Abrol, R. P. Kapil // Biological Sciences. — 1991. — Vol. 57 (2). — P.127–132.
- Bogdanov S. Sugar profiles and conductivity of stingless bee honeys from Venezuela / S. Bogdanov, P. Vit, V. Kilchenmann // Apidologie. — 1996. — Vol. 27 (6). — P.445–450.
- Dabson H. E. Survey of pollen and pollenkitt lipids — chemical cues to flower visitors? / H. E. Dabson // Amer. J. Bot. — 1988. — Vol. 75 (2). — P.170–182.
- Biosynthesis of makisterone A and 20-hydroxyecdysone from labeled sterols by the honey bee, Apis mellifera / F. M. Feldlaufer, E. W. Herbert, J. A. Svoboda, M. J. Thompson // Arch Insect Biochem Physio. — 1986. — Vol. 3. — P.415–421.
- Sterol utilization in honey bees fed a synthetic diet: effects on brood rearing / E. W. Herbert Jr., J. A. Svoboda, M. J. Thompson, H. Shimanuki // J. Insect Physiol. — 1980. — Vol. 26 (5). — P.287–289.
- Major quantitative trait loci affecting honey bee foraging behavior / G. J. Hunt, R. E. Page Jr., M. K. Fondrk, C. J. Dullum // Genetics. — 1995. — Vol. 141 (4). — P.1537–1545.
- Johanson T. S. Feeding honey-bees pollen and pollen substitutes / T. S. Johanson, M. P. Johanson // Bee Word. — 1977. — Vol. 58 (3). — P.105–118.
- Keller I. Pollen nutrition and colony development in honey bees — Part II / I. Keller, P. Fluri, A. Imdorf // Bee word. — 2005. — Vol. 86 (2). — P.27–34.
- Page R. E. J. Genetic determinants of honey bee foraging behavior / R. E. Page Jr., K. D. Waddington, G. J. Hunt, M. K. Fondrk // Animal Behaviour. — 1995. — Vol. 50 (6). — P.1617–1625.
- Analysis of pollen and nectar of Arbutus unedo as a food source for Bombus terrestris (Hymenoptera: Apidae) / P. Rasmont, A. Regali, T. Ings, G. Lognay, E. Baudart, M. Marlier, E. Delcarte, P. Viville, C. Marot, P. Falmagne, J. Verhaeghe, L. Chittka // J. Econ. Entomol. — 2005. — № 98. — P.656–663.
- Comparison of sterols of pollens, honeybee workers, and prepupae from field sites / J. A. Svoboda, E. W. Herbert Jr., W. R. Lusby, M. J. Thompson // Archives of Insect Biochemistry and Physiology. — 1983. — Vol. 1 (1). — P.25–31.
- Utilization and metabolism of dietary sterols in the honey bee and the yellow fever mosquito / J. A. Svoboda, M. J. Thompson, E. W. Herbert Jr., T. J. Shortino // Lipids. — 1982. — Vol. 17. — P.220–225.
- Is non-host pollen suitable for generalist bumblebees? / М. Vanderplanck, S Decleves., N. Roger, C. Decroo, G. Caulier, G. Glauser, P. Gerbaux, G. Lognay // Insect Sci. — 2018. — Vol. 25 (2). — P.259–272.
- How does pollen chemistry impact development and feeding behaviour of polylectic bees? / M. Vanderplanck, R. Moerman, P. Rasmont, G. Lognay, B. Wathelet, R. Wattiez, D. Michez // PLOS ONE. — 2014. — Vol. 9 (1). — P.1–9.
The attractiveness of Maral root for honey bee
Avdeev N. V., PhD Biol. Sc.
Perm Agricultural Research Institute – branch of the Perm Federal Research Center of the Ural Branch of the Russian Academy of Sciences,
614532, Russia, the Perm Territory, Permskiy rayon, selo Lobanovo (village), Kultury str., 12
E-mail: permbee@yandex.ru
The investigation took place at the Perm Agricultural Research Institute on Maral root and honey bees in 2019. The aim was to study the possibility of Maral root pollination by honey bees in the Perm region. The proportion of bees was estimated that pollinated the plants when collecting nectar and pollen. For a nectar to be chosen sugar content and ratio are of great importance. Maral root is a promising medicinal and forage plant whose cultivation is limited by seeds of low quality. Bees were caught at the Maral root flowering time and their pollen pellet was analyzed. The respective literature and microscope slides were used to identify pollen grains. Pollen analysis revealed that Maral root produced insufficient amount of nectar in summer 2019. However, bees actively collected Maral root pollen. The proportion of bees carrying Maral root pollen pellet amounted to 22.5% which was a very high number, considering Maral root growing area (0.6 ha) and presence of other entomophilous plants. It is likely that Maral root pollen contained ecdysteroids — the reason of its high attractiveness for honey bees. Pollination of Maral root by honey bees can become an important element in its seed production technology.
Keywords: honey bee, Maral root, pollen analysis, pollination, ecdysteroid.
References
1. Avdeev N. V. Osobennosti troficheskoy spetsializatsii raznykh ekologicheskikh grupp evro-sibirskoy pchely Apis mellifera mellifera L.: avtoref. dis. … kand. biol. nauk: 03.00.16, 06.02.04. — Moscow, 2006. — 108 p.
2. Alyabeva G. N. Okhrana i ratsionalnoe ispolzovanie endemichnykh rasteniy Kuznetsko-Salairskogo nagorya / G. N. Alyabeva, S. V. Ovsyannikova // Sovremennye naukoemkie tekhnologii. — 2005. — No. 5. — P.51–53.
3. Akhrem A. A. Ekdisteroidy: khimiya i biologicheskaya aktivnost / A. A. Akhrem, N. V. Kovganko. — Minsk: Nauka i tekhnika, 1989. — 327 p.
4. Burmistrov A. N. Medonosnye rasteniya i ikh pyltsa / A. N. Burmistrov, V. A. Nikitina. — Moscow: Rosagropromizdat, 1990. — 192 p.
5. Gaydak M. G. Zhiznedeyatelnost medonosnykh pchel / M. G. Gaydak // Pchela i uley. — Moscow: Kolos, 1969. — P.80–148.
6. Rapontik saflorovidnyy v kulture na Evropeyskom Severo-Vostoke (ekologo-fiziologicheskoe issledovanie) / T. K. Golovko, E. V. Garmash, S. V. Kurenkova, G. N. Tabalenkova, Yu. M. Frolov. — Syktyvkar: Komi nauchnyy tsentr UrO RAN, 1996. — 140 p.
7. Daynen L. Strategiya otsenki roli fitoekdisteroidov kak deterrentov po otnosheniyu k bespozvonochnym fitofagam / L. Daynen // Fiziologiya rasteniy. — 1998. — Vol. 45. — No. 3. — P.347–359.
8. Zherebkin M. V. O nekotorykh osobennostyakh pyltsesobiratelnoy deyatelnosti pchel raznykh porod / M. V. Zherebkin, R. K. Mironova // Vestnik. — 1976. — No. 25.— P.3–61.
9. Zauralov O. A. Rastenie i nektar. Obrazovanie i vydelenie nektara / O. A. Zauralov. — Saratov: Izd-vo Saratovskogo universiteta, 1985. — 180 p.
10. Zauralov O. A. Florospetsializatsiya u medonosnykh pchel / O. A. Zauralov // Zhurnal obshchey biologii. — 1981. — Vol. 42. — No. 6. — P.844–856.
11. Atlas pyltsevykh zeren (Pollen atlas) / I. V. Karpovich, E. S. Drebezgina, E. A. Elovikova, G. I. Legotkina, E. N. Zubova, R. Z. Kuzyaev, R. Kh. Khismatullin. — Ekaterinburg: Uralskiy rabochiy, 2015. — 318 p.
12. Karusevich A. A. Razrabotka metodiki kultivirovaniya i izucheniya osobennostey razvitiya levzei saflorovidnoy v Vitebskoy oblasti / A. A. Karusevich, G. N. Osochuk // Vestnik farmatsii. — 2016. — No. 4 (74). — P.38–44.
13. Raznoobrazie antofilnykh nasekomykh i biokhimicheskie osobennosti nektara introdutsentov roda Serratula / S. V. Pestov, N. I. Filippov, K. G. Ufimtsev, V. V. Volodin, S. O. Volodina, A. G. Dontsov, I. V. Beshley // Teoreticheskaya i prikladnaya ekologiya. — 2017. — No. 3. — P.91–96.
14. Polishchuk V. P. Sbor pyltsy v period medosbora / V. P. Polishchuk // Pchelovodstvo. — 1984. — No. 11. — P.12–13.
15. Ruban G. A. Osobennosti semennoy reproduktsii levzei saflorovidnoy / G. A. Ruban, K. S. Zaynullina // Agrarnaya nauka Evro-Severo-Vostoka. — 2013. — No. 4 (35). — P.22–25.
16. Timofeev N. P. Raspredelenie 20-gidroksiekdizona v strukture biomassy nadzemnoy chasti Rhaponticum carthamoides (Willd.) Iljin v usloviyakh agrotsenoza (Respublika Komi) / N. P. Timofeev, V. V. Volodin, Yu. M. Frolov // Rastitelnye resursy. — 1998. — Vol. 34. — No. 3. — P.63–69.
17. Timofeev N. P. Semennaya reproduktsiya i povrezhdaemost vreditelyami rasteniy s gormonalnoy aktivnostyu nasekomykh / N. P. Timofeev // Ekologo-populyatsionnyy analiz poleznykh rasteniy: introduktsiya, vosproizvodstvo, ispolzovanie. — Syktyvkar: Izd-vo Komi NTs UrO RAN, 2008. — P.192–194.
18. Timofeev N. P. Ekologicheskoe vzaimodeystvie agropopulyatsii ekdisteroidsoderzhashchikh rasteniy s nasekomymi-fitofagami. Soobshchenie 2. Izmenchivost sostava fitoekdisteroidov v argotsenoze i ikh rol v porazhaemosti rasteniy vreditelyami / N. P. Timofeev // Sibirskiy ekologicheskiy zhurnal. — 2009. — No. 6. — P.829–842.
19. Timofeev N. P. Effekt malykh doz ekdisteroidov v pchelovodstve / N. P. Timofeev // Materialy IV mezhdunarodnoy nauchno-proizvodstvennoy konferentsii “Selektsiya, ekologiya, tekhnologiya vozdelyvaniya i pererabotki netraditsionnykh rasteniy”. — Simferopol, 1996. — P.231–232.
20. Ufimtsev K. G. Fitoekdisteroidy — deterrenty nasekomykh-fitofagov / K. G. Ufimtsev, T. I. Shirshova, V. V. Volodin. — Ekaterinburg: Izd-vo Komi NTs UrO RAN, 2009. — 89 p.
21. Ufimtsev K. G. Fitoekdisteroidy kak deterrenty nasekomykh-fitofagov: deystvie rasteniya serpukhi ventsenosnoy Serratula coronata L., produtsenta ekdisteroidov, na egipetskuyu khlopkovuyu sovku Spodoptera littorali (Lepidoptera: Noctuida) / K. G. Ufimtsev, T. I. Shirshova, V. V. Volodin // Uspekhi sovremennoy biologii. — 2009. — Vol. 129. — No. 3. — P.1–15.
22. Frish K. Iz zhizni pchel / K. Frish. — Moscow: Mir, 1980. — 216 p.
23. Abrol D. P. Foraging strategies of honeybees and solitary bees as determined by nectar sugar components / D. P. Abrol, R. P. Kapil // Biological Sciences. — 1991. — Vol. 57 (2). — P.127–132.
24. Bogdanov S. Sugar profiles and conductivity of stingless bee honeys from Venezuela / S. Bogdanov, P. Vit, V. Kilchenmann // Apidologie. — 1996. — Vol. 27 (6). — P.445–450.
25. Dabson H. E. Survey of pollen and pollenkitt lipids — chemical cues to flower visitors? / H. E. Dabson // Amer. J. Bot. — 1988. — Vol. 75 (2). — P.170–182.
26. Biosynthesis of makisterone A and 20-hydroxyecdysone from labeled sterols by the honey bee, Apis mellifera / F. M. Feldlaufer, E. W. Herbert, J. A. Svoboda, M. J. Thompson // Arch Insect Biochem Physio. — 1986. — Vol. 3. — P.415–421.
27. Sterol utilization in honey bees fed a synthetic diet: effects on brood rearing / E. W. Herbert Jr., J. A. Svoboda, M. J. Thompson, H. Shimanuki // J. Insect Physiol. — 1980. — Vol. 26 (5). — P.287–289.
28. Major quantitative trait loci affecting honey bee foraging behavior / G. J. Hunt, R. E. Page Jr., M. K. Fondrk, C. J. Dullum // Genetics. — 1995. — Vol. 141 (4). — P.1537–1545.
29. Johanson T. S. Feeding honey-bees pollen and pollen substitutes / T. S. Johanson, M. P. Johanson // Bee Word. — 1977. — Vol. 58 (3). — P.105–118.
30. Keller I. Pollen nutrition and colony development in honey bees — Part II / I. Keller, P. Fluri, A. Imdorf // Bee word. — 2005. — Vol. 86 (2). — P.27–34.
31. Page R. E. J. Genetic determinants of honey bee foraging behavior / R. E. Page Jr., K. D. Waddington, G. J. Hunt, M. K. Fondrk // Animal Behaviour. — 1995. — Vol. 50 (6). — P.1617–1625.
32. Analysis of pollen and nectar of Arbutus unedo as a food source for Bombus terrestris (Hymenoptera: Apidae) / P. Rasmont, A. Regali, T. Ings, G. Lognay, E. Baudart, M. Marlier, E. Delcarte, P. Viville, C. Marot, P. Falmagne, J. Verhaeghe, L. Chittka // J. Econ. Entomol. — 2005. — No. 98. — P.656–663.
33. Comparison of sterols of pollens, honeybee workers, and prepupae from field sites / J. A. Svoboda, E. W. Herbert Jr., W. R. Lusby, M. J. Thompson // Archives of Insect Biochemistry and Physiology. — 1983. — Vol. 1 (1). — P.25–31.
34. Utilization and metabolism of dietary sterols in the honey bee and the yellow fever mosquito / J. A. Svoboda, M. J. Thompson, E. W. Herbert Jr., T. J. Shortino // Lipids. — 1982. — Vol. 17. — P.220–225.
35. Is non-host pollen suitable for generalist bumblebees? / М. Vanderplanck, S Decleves., N. Roger, C. Decroo, G. Caulier, G. Glauser, P. Gerbaux, G. Lognay // Insect Sci. — 2018. — Vol. 25 (2). — P.259–272.
36. How does pollen chemistry impact development and feeding behaviour of polylectic bees? / M. Vanderplanck, R. Moerman, P. Rasmont, G. Lognay, B. Wathelet, R. Wattiez, D. Michez // PLOS ONE. — 2014. — Vol. 9 (1). — P.1–9.